Liver Fibrosis . Gut-Liver Axis . Bacterial Metabolites . MR1-Restricted Antigen-Presentation . Mucosal-Associated Invariant T Cells . Primary Liver Cells
Liver Immunology
Characterization of mucosal-associated invariant T (MAIT) cells in liver diseases
Mucosal-associated invariant T (MAIT) cells represent the most abundant T cell type in human liver. MAIT cells respond to bacterial metabolites presented by MHC-like molecule MR1 on antigen (Ag)-presenting cells (APCs). In mouse models, activated liver MAIT cells were described as profibrogenic and MAIT cells from patients with liver cirrhosis showed functional alterations consistent with pro-fibrotic functions. Gut dysbiosis has also been implicated in progressive liver fibrosis in fatty liver disease in mice. Since liver MAIT cells respond to bacterial metabolites and exhibit pro-fibrotic properties, they likely also contribute to pathogenesis of non-alcoholic fatty liver disease (NAFLD). In our research, we investigate which cells in the human liver are involved in MAIT cell activation and how their profibrogenic function could be prevented. We also study influence of the gut microbiome on the phenotype of MAIT cells, with an emphasis on a pathogenic role of MAIT cells in fibrosis and NAFLD.
We found that several classes of primary liver cells, including hepatocytes, hepatic stellate cells (HSCs), liver endothelial and biliary epithelial cells, have the capacity to present bacterial Ag to MAIT cells. Liver cells exposed to bacterial Ag precursor had the capacity to generate active Ag endogenously. The identified interaction between HSCs and MAIT cells supports the pro-fibrogenic function of these cells. The finding that activated liver MAIT cells produce large amounts of IL-17, suggests the involvement of this pro-fibrotic cytokine in liver disease pathogenesis. The observed repression of MAIT cell activation by non-stimulatory MR1 ligands creates a therapeutic opportunity to interfere with MAIT cell pro-fibrogenic properties. To better understand the role of MAIT cell stimulation in liver disease, we are currently studying molecular signatures of MAIT cells derived from liver in comparison to colon and blood and we are also correlating MAIT cell phenotypes with the gut microbiome of patients with liver diseases.
Mechanisms of MAIT cell regulation in the liver – physiological functions and pro-fibrotic properties
We assessed the localization of MAIT cells by immunofluorescence staining of cryopreserved human liver tissue and found them to dispersedly localize within the liver parenchyma, thus being in close contact to cells within the sinusoidal compartment (Fig. 1). We have found that distinct primary liver cell types, including hepatocytes, HSCs, liver sinusoidal endothelial cells (LSECs), and biliary epithelial cells (BECs), act as strong drivers of MAIT cell activation, with the potential of increasing liver fibrogenesis (Fig. 2). Presentation occurred in response to both pure synthetic Ag 5-OP-RU (5-(2-oxopropylideneamino)-6-D-ribitylamino-uracil) and bacterial lysate from E. coli. Presentation capacities differed markedly among the investigated liver cell types, with hepatocytes being the most efficient liver-derived APCs (data not shown).
MAIT cell activation was prevented by pre-treating APCs with the non-stimulatory MR1 ligands 6-formylpterin (6-FP) and Acetyl-6-FP and was diminished in the presence of the acetylsalicylic acid derivative 5-formylsalicylic acid (data not shown). These findings create a therapeutic opportunity to prevent pro-fibrogenic properties of MAIT cells. Human polyclonal liver-derived MAIT cells produced large amounts of Interleukin 17 (IL-17), a cytokine with strong pro-fibrotic properties, when interacting with distinct liver-derived APCs, including hepatocytes and HSCs. Moreover, liver cells exposed to an Ag precursor (5-A-RU) had the capacity to generate active Ag 5-OP-RU, as assessed by Ag-presentation assays and mass spectrometry (data not shown).
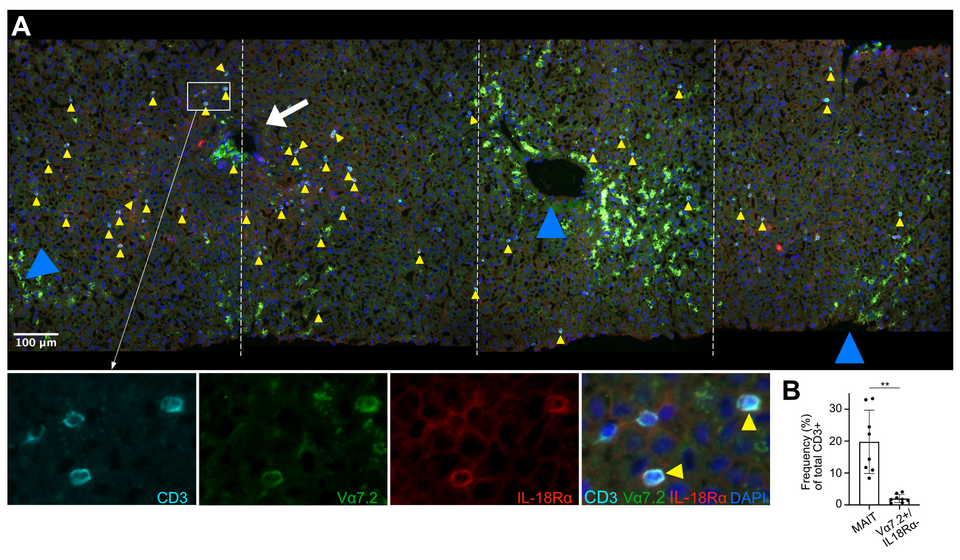
Fig. 1: MAIT cells localize dispersedly to the parenchymal space in healthy human liver.(A) Representative IF analysis of tissue section from a liver biopsy without histopathological abnormalities. Co-localization of CD3, TCR Vα7.2 and IL18-Rα (see higher magnification lower panels) identifies MAIT cells (yellow arrow heads). White arrow and blue arrow heads point at portal field and central veins, respectively. Lower panels also show MAIT cells in proximity of TCR Vα7.2- and IL18-Rα-negative T cells. (B) Percentages of MAIT cells and non-MAIT Vα7.2+ cells versus total CD3+ T cells in healthy human liver (n= 8), assessed as shown in (A). ** P < 0.01, non-paired Welch t-test.
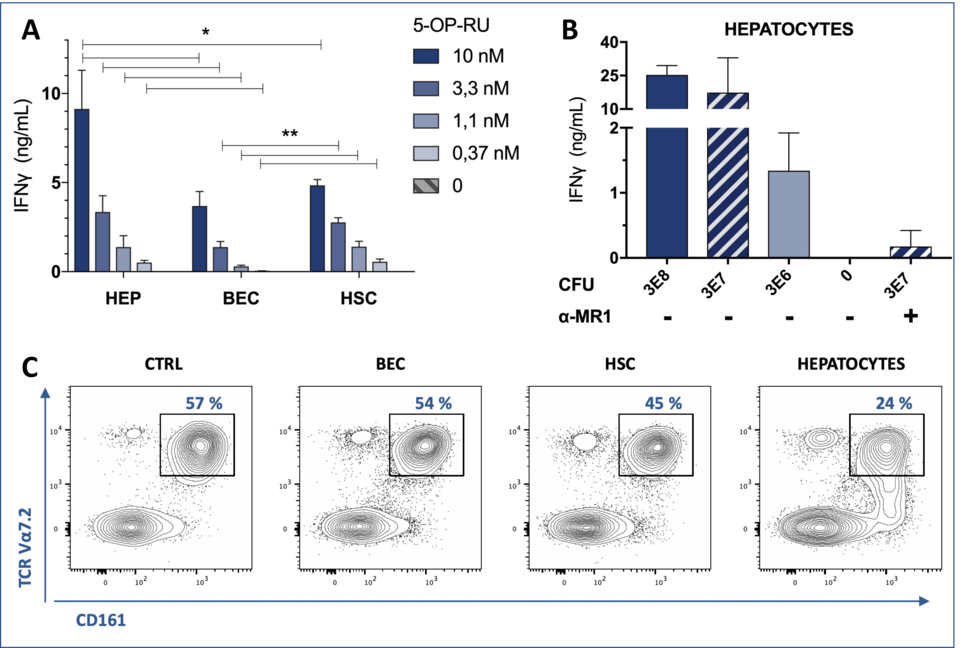
Fig. 2: Robust MAIT cell activation in response to interactions with primary human liver cells. Synthetic antige (Ag) 5-OP-RU (A), and E. coli lysate (B) titration on liver-derived cells, including hepatocytes (HEP), BEC, and HSC, used as Ag-presenting cells (APCs). IFN-γ production by a liver-derived MAIT cell line (A) and blood-derived MAIT cell clone (B) is depicted. MR-1 dependence of the activation process was confirmed by MR1-blocking antibodies. (C) Cell surface staining for MAIT cell markers CD161 and Vα7.2 in response to distinct liver-derived APCs exposed to 5 nM 5-OP-RU. The left panel corresponds to the negative control (CTRL) lacking the APC. Primary blood-derived Vα7.2+ CD161++ cells were enriched by negative selection.