Adult Neurogenesis . Hippocampus . Synaptic Transmission. Neuronal Excitability Dendritic Integration . Calcium Signaling
Cellular Neurophysiology
Synaptic integration of young neurons into the adult hippocampus
In the adult hippocampus new neurons are continuously generated throughout life by adult neural stem cells (Fig. 1). The newly generated young neurons show a number of distinct functional properties, including enhanced excitability, reduced GABAergic inhibition and enhanced synaptic plasticity (Lodge and Bischofberger 2019). Based on these cell biological findings it was concluded that the young neurons are hyperactive and hyperexcitable during learning behavior and memory processing. However, on the behavioral level adult neurogenesis improves learning by increasing the brains capability to distinguish between similar memory items, a process called “pattern separation”. During the last research period (2017–2020) we focussed on glutamatergic and GABAergic circuit analysis, to address this apparent paradox of “hyperactive cells” versus “improved pattern separation”.
Synaptic recruitment of adult-born young granule cells. We studied excitatory glutamatergic synaptic transmission from cortical perforant-path fibers onto newly generated young granule cells (GCs) up to 4 weeks post mitosis (wpm, Li et al. 2017). We found that the young neurons fire action potentials (APs) as early as 2 wpm in response to a small number of active glutamatergic synapses, due to a high synaptic gain. However, due to small dendritic trees and sparse connectivity, neighboring young neurons are activated by different distinct small subsets of afferent fibers with minimal overlap. As the neurons mature, the increase in synapse number is balanced by a gradual decrease in intrinsic excitability. This indicates that the enhanced excitability in young granule cells does not generate hyperexcitability, but instead compensates for sparse synaptic connectivity in developing young neurons. Using paired whole-cell recordings, we could show that AP firing in neighboring young cells is not unspecific, but rather dependent on small non-overlapping populations of afferent input fibers (Fig. 2). This is due to the sparse connectivity combined with the high synaptic gain, generating differential and highly specific spiking output in neighboring cells. Therefore, perforant-path fibers can recruit young neurons in a sparse and orthogonal manner, well suited to support sparse coding during hippocampal information processing.
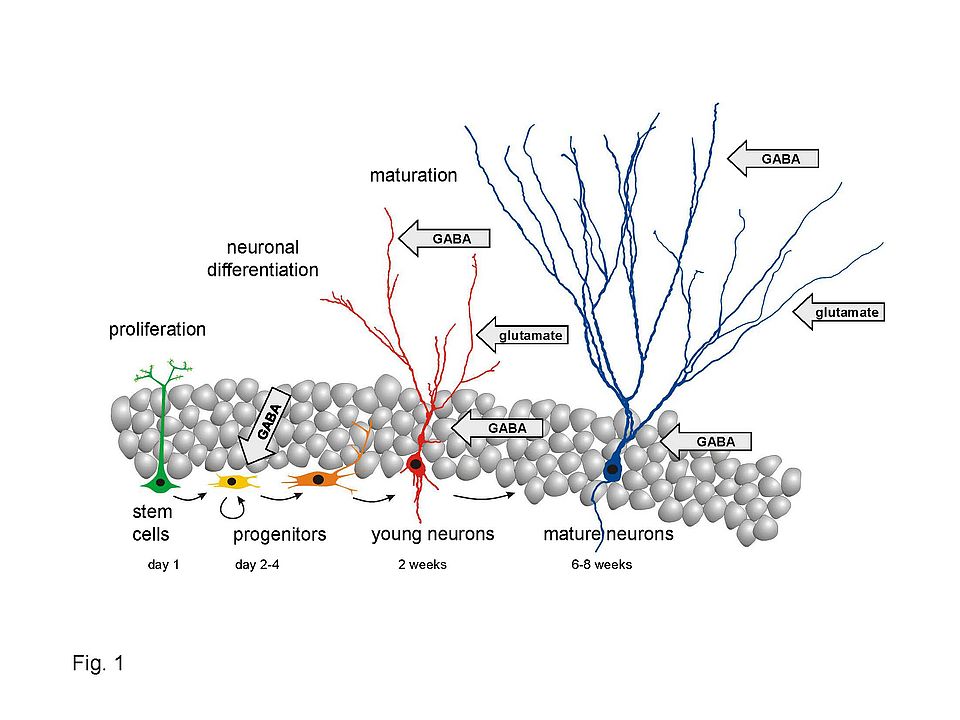
Fig. 1: Adult neurogenesis in the hippocampus. Adult neural stem cells (green) are localized within the subgranular zone of the hippocampus and give rise to transient amplifying cells (yellow). They generate postmitotic neuroblasts which subsequently differentiate into young neurons (red). During the following ~6 weeks, they form new dendrites and several thousand new synaptic connections with glutamatergic neurons as well as with various types of soma- and dendrite-targeting GABAergic interneurons.
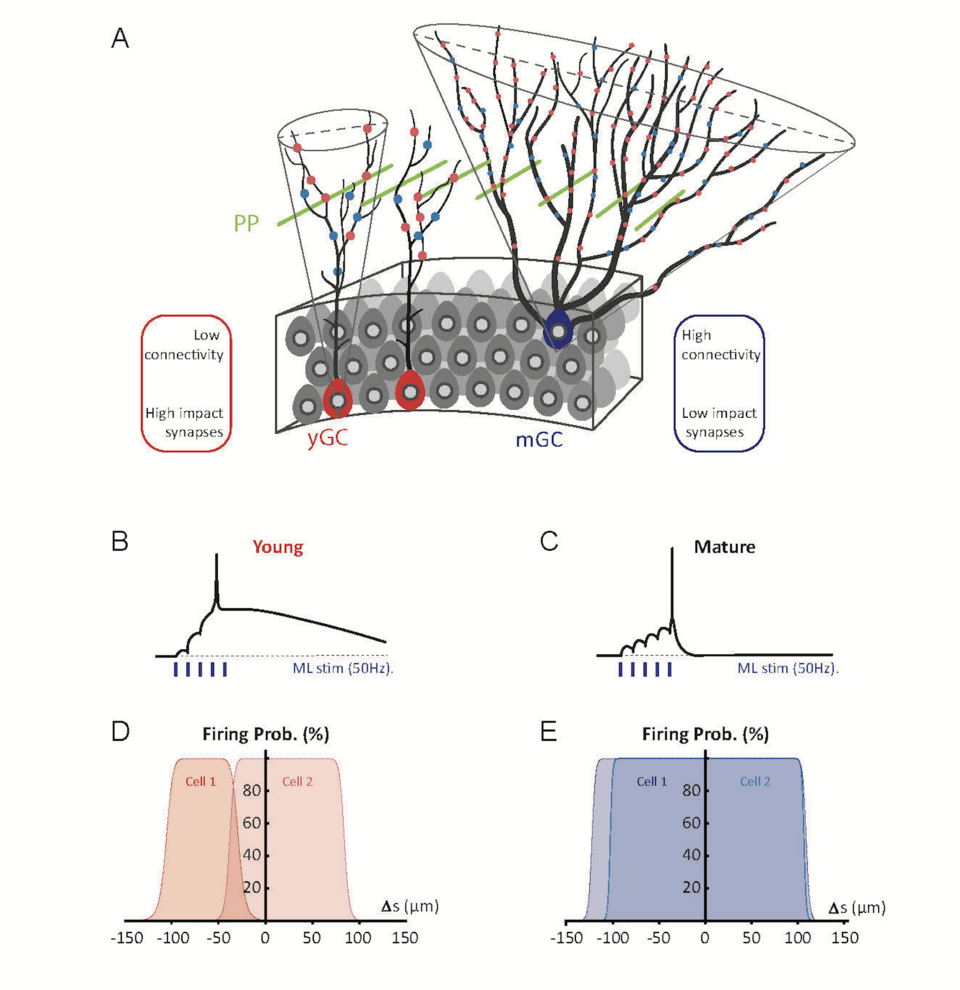
Fig. 2: Low connectivity and high excitability in young granule cells supports orthogonal firing. (A) Schematic representation of the dentate. Young granule cells (shown in red) have smaller dendritic cones (outlined in grey) than mature GCs (blue). Glutamatergic (red circles) and GABAergic (blue circles) synapses are fewer and have a high impact in depolarizing young cells. Perforant path (PP) fibers (green) traverse the molecular layer perpendicular to the dendritic trees of granule cells. Due to the small dendritic cones of young cells, different PP fibers connect with young GCs in a non-overlapping manner. In contrast, in mature GCs glutamatergic and GABAergic synapses are present in large numbers and widely distributed. (B) Despite low numbers, the synaptic connections in young GCs efficiently generate AP firing via efficient temporal summation and non-linear NMDAR-dependent electrogenesis. (C) Mature cells have low intrinsic excitability, but when combined with a large number of inputs from many synapses, are able to reliably fire action potentials. (D,E) The firing probability of two neighboring young (D) or mature (E) cells plotted against the location (Δs) of stimulated afferent fibers. While young GCs are activated by distinct sets of afferent fibers, mature GCs are activated by spatially overlapping perforant path fibers (from Lodge and Bischofberger 2019).